
Fossilien
Lebensweise der Ceratiten
Die Rekonstruktion der Lebensweise ausgestorbener Organismen basiert auf der Auswertung ihrer fossil erhaltenen Reste und ist vom Zeitpunkt des jeweiligen Kenntnisstandes abhängig. Bislang ist es üblich, fossile Cephalopoden mit Außenschale im Analogievergleich mit dem rezenten Nautilus darzustellen. Für diese Methode sprechen gemeinsame Merkmale, wie das planspiral aufgerollte Gehäuse und die innere Zweiteilung in Wohnkammer mit Phragmokon und Sipho. Obwohl die Morphologie des Weichkörpers fossiler ectocochleater Cephalopoden unbekannt ist, übernimmt man allgemein einen Nautilus ähnlichen Organismus mit Greifarmen und Trichter.
Dementsprechend wird auch vorausgesetzt, dass sich diese Tiere einst wie Nautilus frei beweglich in der Wassersäule über dem Boden aufhielten und damit eine schwimmende Lebensweise suggeriert.
Bei dieser Betrachtungsweise fällt allgemein auf, dass häufig die einfachsten physikalischen Bedingungen nicht konsequent in Betracht gezogen werden. Für das funktionelle Verständnis der morphologischen Besonderheiten der Schalencephalopoden ist jedoch die Berücksichtigung der hydrostatischen Gesetze unumgänglich. Das starre Gehäuse mit seinem vorgegebenen Volumen ermöglicht einen exakten Ansatz zur Anwendung der Gesetze der Hydrostatik. Um die Lebensweise von fossilen Cephalopoden mit einer Außenschale nachvollziehen zu können, müssen deshalb vorrangig drei hydrostatische Aspekte, nämlich Auftrieb und Gewicht, die mögliche Schwebefähigkeit sowie die stabile Gehäuseorientierung vorurteilsfrei berücksichtigt werden. Die Arbeit mit diesen Kräften ist ein hervorragendes Mittel, um nicht zutreffende Möglichkeiten mit Sicherheit ausschließen zu können.
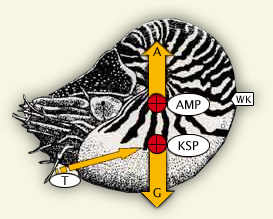
Das Gewicht der von einem Nautilus verdrängten Wassermenge entspricht der nach oben gerichteten Auftriebskraft (A). Diese Kraft setzt im Auftriebsmittelpunkt (AMP) an und bleibt konstant. Das Körpergewicht des Nautilus setzt sich aus der Summe der Teilgewichte von Schale, Weichkörper und Kammerinhalten zusammen und ist veränderlich. Die nach unten gerichtete Gewichtskraft (G) setzt am Körperschwerpunkt (KSP) an. Da beim Schwimmen der KSP immer senkrecht unter dem AMP liegt, wird damit die Gehäusestellung gesetzmäßig vorgegeben. Der mit ca. 135° kurzen Wohnkammer (WK) verdankt Nautilus die günstige Mündungsorientierung; dem relativ großen Abstand von AMP : KSP seine verhältnismäßig stabile Lage. Diese Bedingungen ermöglichen die bestmögliche horizontale Fortbewegung mit einem vom Boden wegführenden Trichterschub.
Auftrieb und Gewicht
Ein Körper, der in eine Flüssigkeit eingetaucht wird, verdrängt eine Flüssigkeitsmenge die seinem Volumen entspricht. Er erzeugt dabei eine nach oben gerichtete Kraft, den Auftrieb. Definitionsgemäß greift diese Kraft im Körpermittelpunkt, dem Auftriebszentrum (AMP) an. Diese Auftriebskraft entspricht dem Produkt aus Volumen und spezifischem Gewicht des Wassers, d.h. dem Gewicht der verdrängten Flüssigkeit. Dabei ist es egal aus welchem Stoff der Körper besteht und ob er in der Wassersäule schwebt, sinkt oder aufsteigt.
Die Auftriebskraft entspricht immer dem Gewicht der verdrängten Flüssigkeit und nicht der Masse des Cephalopoden.
Ein am Boden liegendes, postmortal verfülltes Ceratitengehäuse erzeugt als Steinkern die gleiche Auftriebskraft wie ein lebender Ceratit mit Weichkörper und gasgefülltem Phragmokon gleichen Volumens. Der in der geschlossenen Schale befindliche Phragmokon verdrängt kein Wasser zusätzlich und kann somit auch nicht den Auftrieb beeinflussen.
Im Gegensatz zum unveränderlichen Auftrieb eines starren Körpers ist sein Gewicht veränderbar. Das Gewicht eines Schalencephalopoden setzt sich aus der Summe aller Teilgewichte von Schale, Weichkörper und Kammerinhalten zusammen. Der Angriffspunkt der Gewichtskraft befindet sich im Körperschwerpunkt (KSP). Da dieser gesetzmäßig senkrecht unter dem Auftriebszentrum liegt, wird damit die Richtung der konträr wirkenden Kräfte fixiert und die endgültige Gehäusestellung vorgegeben. Diese Position kann im Schwebezustand vom Organismus nicht beeinflusst werden.
Ausschließlich die Differenz zwischen Auftriebskraft und Gewichtskraft entscheidet darüber, ob ein Körper aufsteigt, schwebt oder sinkt bzw. wie der Ceratitensteinkern am Boden liegen bleibt.
Diese von Archimedes (287-212 v. Chr.) entdeckten Gesetzmäßigkeiten werden inzwischen seit mehr als 2000 Jahren mit Erfolg in Forschung und Praxis angewandt.
Allgemein wird das Aufsteigen eines Körpers doppeldeutig gleichfalls mit Auftrieb bezeichnet, obwohl beide Begriffe inhaltlich nicht identisch sind. Um Irritationen zu vermeiden, ist es deshalb exakter, die Differenz zwischen Auftriebskraft und Gewichtskraft prinzipiell mit "resultierender Auftrieb" zu bezeichnen. Diese resultierende Kraft ist veränderlich und kann ein positives oder negatives Vorzeichen besitzen. Ein sinkender Körper hat logischerweise einen negativen resultierenden Auftrieb.
Schwebegleichgewicht
Die Herstellung eines Schwebezustands ist ein Spezialfall des resultierenden Auftriebs mit dem Wert Null (Auftriebskraft = Gewichtskraft). Das Schwebegleichgewicht stellt eine sehr empfindliche Konstellation dar, die sich kaum genau erreichen und beibehalten lässt. Sie wird deshalb auch vom spezialisierten Nautilus nur näherungsweise aufrechterhalten. Weil die fossilen Schalencephalopoden wie Nautilus auch über einen Kammerapparat mit Sipho verfügen, wird dessen Fähigkeit, damit in Grenzen ein Schwebegleichgewicht regulieren zu können, einfach bei den fossilen Formen vorausgesetzt.

Die Bedeutung der Wohnkammerlänge für die stabile Mündungsorientierung
a: Ceratit (Schema) mit 180° langer Wohnkammer ohne Ballastwasser in Schwebestellung. Kammern des Phragmokons im Vakuum und damit mit geringster Gewichtskraft. Obwohl die Kraftrichtung des Trichterschubes schon leicht zum Boden gerichtet ist, wäre sowohl ein Schwebegleichgewicht als auch eine horizontale Lokomotion trotz großer Instabilität theoretisch noch denkbar. Da sich ohne Ballastwasser jedoch keine Gleichgewichtslage austarieren lässt, entfällt diese Möglichkeit für Ceratites.
b: Ceratit (Schema) mit 180° langer Wohnkammer und Ballastwasser (B) in Schwebestellung. Das Wasser in den zwei letzten Kammern wirkt mit seinem Zusatzgewicht wie ein verlängerter Weichkörper. Die damit faktisch auf >200° verlängerte Wohnkammer bewirkt einerseits mit dem Zusammenrücken von Körperschwerpunkt (KSP) und Auftriebsmittelpunkt (AMP) einen Stabilitätsverlust (nur noch ca. 20% der Stabilität von Nautilus) und anderseits mit dem Anheben der Gehäusemündung einen stets zum Boden gerichteten Trichterschub (T). Zwar wäre jetzt theoretisch das Austarieren einer Gleichgewichtslage möglich, eine horizontale Fortbewegung mit Trichterschub, bei dem jeder Impuls das Gehäuse zum Boden drücken würde, jedoch nicht mehr denkbar.
c: Ammonitengehäuse (Schema) mit 300° langer Wohnkammer ohne Ballastwasser oder Ceratit (Schema) mit 180° langer Wohnkammer mit bis auf 300° aufgefülltem Ballastwasser in Schwebestellung. Auftriebsmittelpunkt (AMP) und Körperschwerpunkt (KSP) liegen unmittelbar übereinander und die Gehäusemündung ist maximal nach oben gewandert. In dieser Situation ist das Gehäuse völlig instabil und würde vom geringsten Trichterschub (T) nur noch wie ein Windrad rotierend bewegt.
Stabile Gehäuseorientierung
Da Körpermittelpunkt (AMP) und Körperschwerpunkt (KSP) im Schwebegleichgewicht immer senkrecht übereinander liegen, wird damit die Gehäusestellung mit der Mündungsorientierung gesetzmäßig vorgegeben. Die Stabilität eines eingerollten Gehäuses ist im Schwebegleichgewicht vom Abstand dieser Punkte abhängig und verringert sich kontinuierlich mit der Verlängerung der Wohnkammer. Dabei gilt der Grundsatz, je enger beide Punkte zusammenrücken, um so instabiler wird das System. Die konsequente Berücksichtigung dieser Gesetzmäßigkeiten ist entscheidend für die Beurteilung der Möglichkeiten der Lokomotion (= Ortsveränderung). Nautilus nutzt die rückwärtig gerichtete Energie des aus dem Trichter ausgestoßenen Atemwassers zur horizontalen Fortbewegung. Obwohl bei ihm Körpermittelpunkt und Körperschwerpunkt den für Schalencephalopoden optimalen Abstand haben, bewirkt jeder Rückstoß noch ein Drehmoment, das sich in einer Schaukelbewegung bemerkbar macht. Schnelle horizontale Ortswechsel sind bei dieser Konstruktion prinzipiell limitiert. Jedes weitere Zusammenrücken beider Kraftansatzpunkte macht diese Art der Fortbewegung uneffektiver und schließlich unmöglich (siehe Schema a-c). Abbildungen fossiler Schalencephalopoden in Lebensstellung zeigen immer wieder, dass diese vorgegebene Mündungsorientierung und die davon abhängige Instabilität bei Rekonstruktionsversuchen nicht berücksichtigt wird.
Hydrostatische Gesetze und die Germanischen Ceratiten
Die Gehäuse-Morphologie der Ceratiten des Oberen Muschelkalks ist im Verlaufe ihrer phylogenetischen Entwicklung vielen Veränderungen unterworfen. Lediglich die Länge der Wohnkammer bleibt ab der sequens/pulcher-Zone mit ca. 180° konstant. In ihr befindet sich der Weichkörper, der mit seinem spezifischen Gewicht (g ~ 1,056) die Lage des Körperschwerpunktes entscheidend beeinflusst. Wie das Schema zeigt, wäre Ceratites mit dieser vorgegebenen Gehäuseorientierung und ohne Ballast theoretisch durchaus schwebe- und bedingt schwimmfähig. Da jedoch Auftriebsmittelpunkt (AMP) und Körperschwerpunkt (KSP) bereits sehr dicht beieinander liegen, wird dadurch die Gehäuselage hochgradig instabil und eine mittels Trichterschub erreichbare Horizontalbeschleunigung minimiert. Sie eignet sich weder zum Beutefang noch zur Fluchtmöglichkeit vor Fressfeinden.
Die folgenden Befunde des fossilen Materials werden aus biomechanischer Sicht zeigen, ob sich Ceratiten wirklich frei in der Wassersäule bewegt haben können.
Rückschlüsse aus echter Epökie von Placunopsis auf Ceratites
Eine objektive Argumentation kann nur indirekt über Rückschlüsse auf Reaktionen des Organismus erfolgen, die als Information auf dem Steinkern erhalten sein müssen. Als eine praktikable Möglichkeit erweist sich der häufig zu Lebzeiten des Ceratiten erfolgte Aufwuchs durch die Muschel Placunopsis ostracina. Er wird als echte Epökie bezeichnet und hatte unzweifelhaft eine zusätzliche Gewichtserhöhung zur Folge. Ein Ceratiten-Organismus der ständig sein Schwebegleichgewicht austarieren musste, war bei Epökie gezwungen, auf die zusätzliche Gewichtskraft mit Gegenmaßnahmen zu reagieren. Dafür blieben dem Ceratiten zwei Möglichkeiten zur Verringerung der Gewichtskraft und eine zur Vergrößerung der Auftriebskraft.
Das einfachste Mittel zur Verringerung des Gesamtgewichtes ergibt sich durch Abpumpen von Ballastwasser aus dem Phragmokon.
Eine derartige Reaktion ist auf einem Steinkern nicht nachzuweisen. Allerdings wäre aufgrund der ohnehin nur geringen theoretischen Ballastreserve (s. Schema) damit keine dauerhafte Regulierung erreicht und könnte nur als kurzzeitige Notlösung in Betracht gezogen werden. Da echte Epökie immer eine zusätzliche Belastung auf Lebenszeit darstellt, ist diese Maßnahme auszuschließen.
Eine weitere Möglichkeit wäre der Bau zusätzlicher Kammern der jedoch eine Verkürzung der Wohnkammerlänge zur Folge hätte.
Diese Form des Gewichtsausgleichs ist von zwei abnormal gewachsenen Nautilus pompilius (REIN 2000) bei Aquarienhaltung bekannt. An Stelle eines normalen Gehäusewachstums wurden von den Individuen an der Gehäusemündung nur noch überdimensionierte pathologische Kalkwülste ausgeschieden. Da es zu keiner weiteren Volumenzunahme kommt, wird die Auftriebskraft nicht ausgleichend mit erhöht. Zur Erhaltung des Schwebegleichgewichts musste diese einseitige Gewichtszunahme mit dem Bau zusätzlicher Kammern ausgeglichen werden. Die dadurch immer kürzer werdende Wohnkammer führte unweigerlich zum Tod des Tieres.
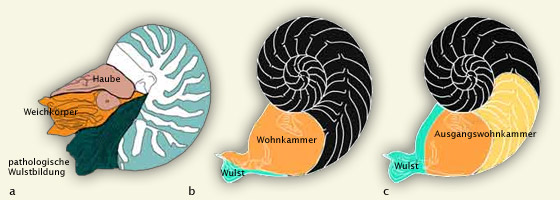
Nautilus und seine begrenzten Möglichkeiten zur Regelung des Schwebegleichgewichtes
a: Nach Aquarienhaltung gestorbener Nautilus;
b: Medianschnitt durch das Gehäuse des gleichen Individuum
c: Rekonstruktion der prämortalen Anpassungsprozesse durch den Weichkörper
Außer der pathologischen Wulstbildung an der Gehäusemündung erfolgte mit Beginn der Aquarienhaltung kein Längenwachstum mehr. Auf das hydrostatische Kräfteverhältnis bezogen bedeutet die fehlende Volumenzunahme eine einseitige Erhöhung der Gewichtskraft. Am Medianschnitt wird deutlich, dass das gestörte Schwebegleichgewicht mit dem zusätzlichen Bau von >6 Kammern vorübergehend austariert werden konnte. Allerdings hatte diese gewichtsmindernde Methode unweigerlich den Tod des Individuums zur Folge, da der Weichkörper faktisch aus der immer kürzer werdenden Wohnkammer geschoben wurde. Nautilus ist aufgrund der hydrostatischen Gesetze nicht befähigt, einseitige Erhöhungen der Gewichtskraft auf Dauer zu kompensieren.
Aus biomechanischer Sicht ist dieses Verhaltensmuster ohne Einschränkung auf alle Cephalopoden mit Außengehäuse übertragbar. Bei einer nektonischen Lebensweise von Ceratites hatte die Gewichtszunahme durch den Aufwuchs der Epöken den gleichen Effekt. Die mit Placunopsis bewachsenen Ceratitensteinkerne müssten in diesem Fall eine zwar nicht pathologisch deformierte, aber deutlich verkürzte Wohnkammer aufweisen, die gleichzeitig den vorzeitigen Tod des Individuums dokumentiert.
 Ceratites weyeri, DE=240 mm, Original REIN (1997), echte Epökie mit >150 Placunopsis ostracina.
Ceratites weyeri, DE=240 mm, Original REIN (1997), echte Epökie mit >150 Placunopsis ostracina.
Etwa 20% der germanischen Ceratiten wurden zu Lebzeiten von Epöken besiedelt. Trotz einseitiger Zunahme der Gewichtskraft sind keine gewichtsvermindernden Reaktionen nachzuweisen. Diese fehlenden Anpassungsmerkmale und die große Überlebensdauer mit dem Zusatzgewicht sind ein Beweis dafür, dass Ceratiten kein Schwebegleichgewicht austarieren mussten.
Erhöhung der Auftriebskraft durch Querschnittsvergrößerung an der Gehäusemündung
Die mit einer Querschnittsvergrößerung verbundene Zunahme der Auftriebskraft konnte die Gewichtserhöhung durch Epökie problemlos ausgleichen. Zudem garantierte ein derartiges Verhalten gleichzeitig das Überleben des Individuums. Der Befund einer Volumenänderung des Gehäuses als Reaktion auf die Gewichtszunahme durch Placunopsis Bewuchs wäre somit ein eindeutiger Beweis für eine nektonische Lebensweise der Ceratiten.
Die Sichtung umfangreichen Beleg-Materials mit nachweislich echter Epökie erfolgte mit der Zielstellung, etwaige diesbezügliche morphologische Veränderungen des Gehäusebaues nachzuweisen. Das Ergebnis ist eindeutig. Einige Individuen tolerierten mehr als 200 Placunopsis problemlos über große Zeiträume auf ihrem Gehäuse (REIN 1996, 1997, 1999). Trotz dieser beträchtlichen Gewichtserhöhung war in keinem Falle eine Tendenz zum Ausgleich des Mehrgewichts zu beobachten.
Das vollständige Fehlen derartiger Anpassungen beweist nach dem Gesetz des Archimedes, dass die Gehäuse nicht im Schwebegleichgewicht gehalten wurden.
Zusätzlich wird mit den biomechanischen Befunden offengelegt, dass die Fähigkeit zur horizontalen Lokomotion durch die Wohnkammerlänge limitiert wird. Das Ceratitengehäuse war demnach zur horizontalen Lokomotion mittels Trichterschub funktionell nicht befähigt.
Bei konsequenter Berücksichtigung hydrostatischer Gesetzmäßigkeiten kann deshalb geschlussfolgert werden, dass Ceratiten keine Schwimmer waren und zur Fortbewegung den ständigen Bodenkontakt brauchten.